一方で、AMPキナーゼ(AMPK)は細胞に普遍的に存在し、細胞内エネルギー状態の監視及びその調節を行っていることから、糖・脂質代謝の領域において治療の標的分子として注目されています。しかしながら、これまでに骨芽細胞でのAMPKの役割についての報告はありませんでした。
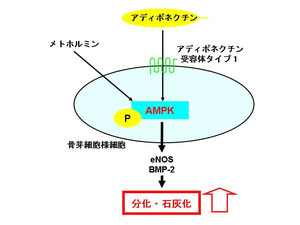
我々はこれまでにアディポネクチンが骨芽細胞においてAMPKを活性化させることにより骨芽細胞の分化、石灰化を促進することを報告し、骨芽細胞におけるアディポネクチンシグナルがAMPKを介していることを世界に先駆けて報告いたしました。
また、我々は2型糖尿病治療薬であるメトホルミンが骨芽細胞においてAMPK活性化を介して骨形成促進因子であるeNOS、BMP-2の発現を誘導し、分化、石灰化を増強することを報告しております。従って、AMPKを活性化しうる薬剤が骨形成を促進し、骨代謝を改善しうる可能性が考えられます。
AMPキナーゼ活性化はメバロン酸経路阻害により骨芽細胞分化を促進する
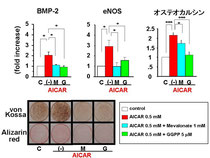
マウス骨芽細胞株であるMC3T3-E1細胞を用いて、AMPK活性化による骨芽細胞の分化、石灰化促進機序をAMPK活性剤であるAICARを用いて検討しました。
AICARはBMP-2、eNOSおよび分化マーカーであるオステオカルシン発現を誘導し、石灰化促進作用を示しました。メバロン酸経路の中間代謝産物であるメバロン酸あるいはグラニルグラニルピロリン酸をAICARと同時投与することによりメバロン酸経路の抑制を解除すると、AICARの作用は消失しました。このことからAICARによるAMPK活性化はメバロン酸経路の阻害によりBMP-2、eNOS、オステオカルシン発現にくわえ石灰化促進作用を発揮していることが明らかとなりました。
また、NOS阻害剤であるL-NAME、あるいはBMPアンタゴニストであるnogginをAICARと同時投与することによりAICARの作用は消失したことから、骨芽細胞におけるAMPK活性化はメバロン酸経路の抑制に引き続くeNOS、BMP-2発現増強を介して、石灰化を促進することが示唆されました。
さらに我々はAICARはAkt、ERKのリン酸化を促進し、ERK活性阻害剤(PD98059)によりAICARのeNOS、BMP-2発現増強作用は抑制されることを示しました。
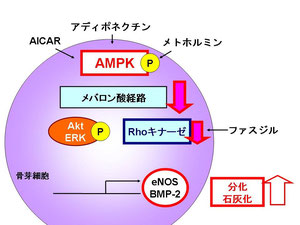
一方で、Rhoキナーゼはメバロン酸経路の下流に存在するが、AICARはRhoキナーゼ活性を有意に抑制し、Rhoキナーゼの特異的阻害剤であり動脈硬化治療薬として注目されているファスジルはAICARと同様にeNOS、BMP-2、オステオカルシン発現を増強し、同時に石灰化も促進しました。
このことにより、AMPK活性化によるメバロン酸経路阻害とファスジルによるRhoキナーゼの直接の阻害は糖・脂質代謝の治療のみならず、新たな骨粗鬆症の治療手段となりうる可能性が示唆されました。
AMPK活性化はグルココルチコイドによる骨芽細胞分化障害を解除する
臨床の場においてグルココルチコイド(GC)はアレルギー疾患や血液疾患を始め様々な疾患で使用される薬剤です。一方で、GC骨粗鬆症(GIO)はGC投与による副作用として最も注意すべき疾患であり、GC投与後の脆弱性骨折を予防することは臨床的に非常に重要であります。GIOの発症機序としてGCによる骨芽細胞のアポトーシスや分化、石灰化の低下が重要であると考えられているが、未だ不明な点が多いのが現状です。
これまでにGCの骨芽細胞分化阻害作用の一つとしてBMPシグナルの低下、Runx2発現の低下が報告されており、我々は以前にBMPシグナルの低下にはBMP-2アンタゴニストであるFollistatin、Danが関与していることを報告しています。
我々はAMPK活性化、Rhoキナーゼ阻害がともにBMP-2シグナルを増強することから、GCによる骨芽細胞分化抑制に対するAICAR、ファスジルに加えコレステロール合成阻害剤であるスタチンの影響を検討しました。
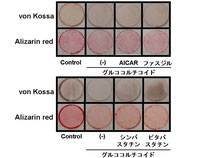
さらに、GC投与によるBMPアンタゴニスト(Follistatin、Dan)の発現増強作用を有意に抑制した。 右図に示すように石灰化染色においてもAICAR、ファスジル、スタチンはいずれもGCによる石灰化抑制作用を解除した。
今後明らかにすること
これまでに我々はAMPKの骨芽細胞分化における制御機構について世界に先駆け報告してきており、その後他のグループからも我々の結果を追従する形で様々なデータが報告されてきています。脂質異常症治療薬であるベザフィブラートもAMPK活性化を介したeNOS、BMP-2発現作用があることが報告されており、またAMPK欠損マウスでは骨量が低下することが確認されています。
1)in vivoにおけるAMPKの骨における役割の検討
上記のようにAMPK欠損マウスにおける骨量低下が報告されていますが、AMPKは全身にユビキタスに存在しており、様々なホルモンのkey moleculeとして働いていることから、実際に骨における役割を検討する上で骨特異的なAMPK遺伝子改変マウスでの検討が必要であると考えています。また、骨粗鬆症モデルマウスにおいて実際にAMPK活性化が骨量低下を改善するかなどについても検討が必要であると考えています。また、我々はGCによる骨芽細胞分化抑制作用へのAMPK活性化の影響をin vitroにて検討しましたが、未だin vivoにおける検討は報告されていません。従って、我々のin vitroでの知見を確認するためにはin vivoにおける検討が重要であると考えています。
2)AMPKのシグナル伝達経路について
近年、他のグループからAMPK活性化はBMP-2シグナルの下流であるSmad1/5/8のリン酸化、Dlx5、Runx2発現を増強することが報告されています。従って、AMPKはBMP-2そのものの発現を増強するのみならずその下流のシグナルを増強することにより骨芽細胞分化促進に働いている可能性があります。また、近年ではBMPシグナルとWntシグナルには関連性があることが報告されてきており、さらなるAMPKシグナル伝達についての検討が必要であると考えています。
3)MAPKとの関係性について
我々の検討でもAMPK活性化はERK活性化を誘導することが確認されています。骨芽細胞におけるAMPKと他のシグナル伝達についての検討は未だ不明な状態です。この点についても今後明らかにする必要があると考えています。
4)AMPKの破骨細胞、骨細胞での役割
AMPKのin vivo、臨床での応用を検討する上で骨芽細胞のみならず他の細胞への影響も当然検討する必要があります。
5)他のホルモンや薬剤のAMPKへの影響
前述したようにAMPKは様々なホルモン作用においても重要な因子であることが分かってきています。従って、骨に重要であるホルモン(PTHやカルシトニン、IGF-Iなど)がAMPKシグナルと関連している可能性も考えられます。